میکروب شناسی پزشکی
مقدمه و تاریخچه
علم میکروب شناسی با اختراع میکروسکوپ در قرن هفدهم توسط آنتونی وان لیون هوک ( 1632 –1723 ) در هلند آغاز گردید. او با مشاهدات خود بر روی آب برکه، بزاق، خلط و... برای اولین بار موفق به مشاهده و کشف دنیای میکروارگانیسم ها شد. او نتایج مشاهدات خود را به انجمن سلطنتی انگلستان گزارش نمود. دوران شکوفایی میکروب شناسی با فعالیت های علمی پاستور (1823 - 1895) ، شیمیدان فرانسوی بر روی پدیده تخمیر آغاز شد. پاستور را پدر علم میکروب شناسی نامیده اند. او نظریه تولید خود به خودی موجودات زنده را رد کرد و ثابت کرد که تخمیر یک فعالیت بیولوژیک و نه صرفا شیمیایی است و توسط بعضی از انواع میکروارگانیسم ها ایجاد می شود. او نقش میکروارگانیسم ها در ایجاد بیماری و فساد مواد غذایی را نشان داد و برای کاهش میکروارگانیسم ها در مواد غذایی روشی را پیشنهاد داد که بعد ها پاستوریزاسیون نام گرفت. پاستور واکسیناسیون را که به شکل تجربی از زمان ادوارد جنر 1749 -) 1823) وجود داشت بر یک مبنای علمی استوار ساخت و برای اولین بار واکسن هایی علیه سیاه زخم و هاری تولید کرد.

در همین دوران یک پزشک آلمانی به نام رابرت کخ (1843 –1910) به میکروب شناسی علاقه مند شد. از مهمترین فعالیت های علمی کخ معرفی اصول چهارگانه ای است که برای تایید بیماری زا بودن یک میکروب مشخص الزامی است. کخ بهمراه همکاران خود موفق شد روش های آزمایشگاهی میکروب شناسی را بهبود بخشد. برای مثال معرفی محیط های کشت جامد با استفاده از نوعی پلی ساکارید دریایی به نام آگار و نیز ابداع ظروف پتری (پتری دیش) از ابتکارات تیم تحقیقاتی او بود. رابرت کخ باکتری های مولد سیاه زخم، وبا و نیز باکتری مولد سل را کشف کرد و بدین سبب در 1905 موفق به کسب جایزه نوبل در پزشکی شد. رابرت کخ را یکی از پایه گذاران اصلی علم میکروب شناسی می شناسند. به دلیل حجم گسترده ی فعالیت های میکروب شناسی و کشف بسیاری از میکروارگانیسم ها در دوران پاستور و کخ، این دوران (1857 - 1914) به نام عصر طلایی میکروب شناسی نامیده شده است. در 1928 الکساندر فلمینگ (1881 –1955)، پزشک انگلیسی پنی سیلین را کشف کرد. این آنتی بیوتیک سپس در سال 1943 به عنوان اولین آنتی بیوتیک به تولید انبوه رسید و در درمان سربازان در اواخر جنگ جهانی دوم بکار گرفته شد و بدین ترتیب عصر آنتی بیوتیک ها آغاز گردید..............
بر روی ادامه مطلب کلیک کنید
طبقه بندی باکتری ها
نام علمی باکتری ها همچون نام علمی دیگر موجودات زنده دو قسمتی است. ابتدا نام جنس (Genus) و به دنبال آن نام گونه (Species) می آید. در زبان انگلیسی حرف اول نام جنس همیشه بزرگ و حرف اول نام گونه کوچک نوشته می شود. نام علمی باکتری ها همچنین در متون انگلیس کج (ایتالیک) نوشته می شود (برای مثال: Echerchia coli ).
باکتری ها اساساً بر اساس تفاوت های مورفولوژیکی و بیو شیمیایی- متابولیکی تقسیم بندی می شوند، اما در عین حال ممکن است از ویژگی های ایمونولوژیک (سرولوژی) و نیز ژنتیکی در طبقه بندی آنها استفاده شود. در میکروب شناسی پزشکی گاهی باکتری ها بر حسب اندامی که محل اصلی عفونت زایی آن هاست طبقه بندی می شوند.
شکل باکتری ها
باکتری های تک سلولی ساده هستند و به اشکال کروی یا کوکسی، میله ای یا باسیل، باسیل کوتاه یا کوکوباسیل، باسیل خمیده یا ویبریون و مارپیچی یا اسپریل وجود دارند. اشکال اسپریل به شکل ویرگول یا s-شکل یا فنری شکل هستند. معدودی از باکتری ها هم که فاقد دیواره هستند شکل ثابتی ندارند (مثل ژل) و پلئومورف خوانده می شوند.
باکتری پس از تکثیر با تجمع در کنار هم الگوهای پیچیده ای مثل کوکسی های دوتایی(دیپلوکوک)، کوکسی های زنجیره ای، (استرپتوکوک)، کوکسی های خوشه ای (استافیلوکوک)، باسیل های زنجیره ای (استرپتو باسیل)، رشته ای (فیلامنتوس) و یا باکتری های منفرد را ایجاد می کند.

شکل1-1-2) اشکال مختلف باکتری ها
اندازه ی باکتری ها
اندازه ی کوکوس ها معمولا بین 75/0 تا 5/1 میکرومتر است. باسیل ها از نظر اندازه متنوع هستند و ممکن است از 5 تا 8 میکرومتر طول و 1 تا 5/1 میکرومتر عرض (مانند باسیل مولد سیاه زخم) تا باسیل های کوچک، مانند هموفیلوس ها که 75/0 تا 5/1 میکرومتر طول و 2/0 تا 3/0 میکرومتر عرض دارند دیده شوند. یاکتری های مارپیچی ممکن است به طول 5 تا 50 میکرومتر و ضخامت 1/0 تا 5/0 میکرومتر باشند.
ساختمان واجزاء ساختمانی باکتری ها
باکتری ها از نظر تکاملی قدیمی ترین گروه موجودات زنده هستند و قدمت آنها به سه بیلیون سال پیش می رسد. به همین دلیل از یک ساختار سلولی ساده برخوردارند که به آن پروکاریوت می گویند. در مقابل سلول های دیگر موجودات زنده تکامل یافته تر هستند و یوکاریوت نام دارند. اندامک هایی که در سلول های یوکاریوتیک موجود است و عمدتا شامل شبکه آندوپلاسمیک، دستگاه گلژی، میتوکندری، لیزوزوم، کلروپلاست (در گیاهان) و واکوئول می باشد در باکتری ها دیده نمی شود. اجزای یک سلول پروکاریوتیک در زیر بیان می شود:
- سیتوپلاسم و اجزای سیتوپلاسمی:
سیتوپلاسم قسمت اصلی سلول باکتری است. زیرا مرکز فعل و انفعالات حیاتی باکتری است. ماتریکس سیتوپلاسم از یک سیستم کلوییدی تشکیل شده است که در آن آب و مواد آلی و غیر آلی از قبیل اسید ریبونوکلئیک، فسفولیپید،گلوسید، پروتئین،ریبونوکلئات منیزیوم و یون های معدنی وجود دارد. تعدادی اندامک در ماتریکس شناورند که شامل هسته ،ریبوزوم ها و گرانول ها هستند. مزوزوم ها نیز که از فرورفتگی پرده سیتوپلاسمی به داخل سیتوپلاسم حاصل می شوند در پیرامون سیتوپلاسم دیده می شود. مزوزوم ها در باکتری های گرم مثبت بزرگتر از گرم منفی ها هستند.
ریبوزوم ها(Ribosomes) . ریبوزوم ها دانه های سیتوپلاسمی به قطر 10 تا 20 نانومتر اند که حاوی 40% پروتئین و 60% RNA می باشند. ریبوزوم ها مرکز پروتئین سازی سلول محسوب می شود. محققان بوسیله میکروسکوپ الکترونی توانسته اند ده ها هزار ریبوزوم در سیتوپلاسم باکتری مشاهده کنند. در باکتری های در حال فعالیت تعدادی ریبوزوم توسط یک رشته RNA به یکدیگر متصل می شوند و ایجاد پلی زوم می نمایند. باکتری ها دارای ریبوزوم های کوچکتر(70S) از سلول های یوکاریوت ((80S هستند. ریبوزوم های باکتریایی از دو ساب یونیت (50S) و (30S) ساخته شده اند .این اعداد مربوط به سرعت رسوب گذاری آنها (Sedimentation) است. آنتی بیوتیک هایی مثل اریترومایسین و تتراسایکلین برای حمله به باکتری، ریبوزوم ها را هدف قرار می دهند. اریترومایسین به ساب یونیت 50S اتصال می یابد و پروتئین سازی را مختل می کند، ولی تتراسایکلین سنتز پروتئین را در ساب یونیت 30S مهار می کند .
گرانول ها یا دانه ها ی سیتوپلاسمی(Granules) . در بسیاری از باکتری ها ذرات کروی در سیتوپلاسم مشاهده می شود که نقش انبار سلولی در ذخیره ی مواد غذایی را به عهده دارند. این اندامک ها جزء لازم ساختمان باکتری است. زمانی که باکتری مواد غذایی در دسترسی دارند اندامک ها به حداکثر اندازه می رسند و زمانی که باکتری با کمبود مواد غذایی مواجه است گرانول ها به حداقل اندازه خود می رسند و یا حتی ناپدید می شوند. هر نوع گرانول ماده غذایی خاصی را ذخیره می کنند. به عنوان مثال، بسیاری از باکتری های دهان گرانول های حاوی گلیکوژن دارند که گرانول های گلوسیدی نامیده می شوند. برخی از باکتری های ساکن در چشمه های آب گرم دارای گرانول های گوگردی دارند. باکتری های چربی دوست نیز غالبا گرانول های لیپیدی دارند. بعضی باکتری ها حاوی چندین نوع گرانول هستند.
هسته (Nucleous). هسته مرکز کنترل باکتری و حاوی اطلاعات ژنتیکی است و هستک و غشای هسته ندارند. در واقع در جایگاه هسته فقط یک مولکول اسید دزوکسی ریبونوکلئیک وجود دارد که کروموزوم نام دارد. بنابراین هر باکتری فقط یک کروموزوم دارد. این مولکول دایره ای شکل است اما بر روی خود پیچ خورده و به شکل کلاف درامده است. طول آن درحدود یک میلیمتر و قطر ان 20 تا 60 آنگستروم است. DNA از طریق مزوزوم به غشای سیتوپلاسمی متصل است. باکتری ممکن است همچنین در سیتوپلاسم خود حاوی حلقه ها ی کوچکی از DNA دو رشته ای باشد که پلاسمید گفته می شود. پلاسمید ها حاوی اطلاعات غیر ضروری، اما سودمند برای بقای باکتری هستند. بسیاری از ژن های مقاومت به آنتی بیوتیک ها در پلاسمید ها جای گرفته اند.
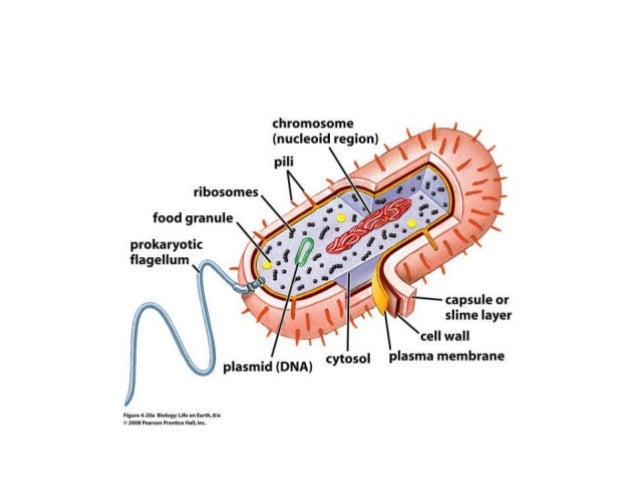
شکل 2-1-2) ساختمان یک سلول باکتری
2. غشای سیتوپلاسمی:
داخلی ترین لایه در جدار یک سلول باکتری، غشای سیتوپلاسمی است. سطح خارجی غشای سیتوپلاسمی را در قریب به اتفاق باکتری ها دیواره سلولی می پوشاند. در بعضی از باکتری ها ممکن است در سطح خارجی دیواره، لایه سومی به نام کپسول دیده شود.
غشای سلولی/ سیتوپلاسمی (Cell/cytoplasmic membrane)
ساختمان فسفو لیپیدی و دو لایه ی غشای سلولی در باکتری ها بسیار به غشای سلولی یوکاریوت ها شبیه است. در عین حال پروتئین غشاء سیتوپلاسمی در باکتری ها از سلول های پستانداران زیادتراست. موادشیمیایی تشکیل دهنده ی پرده ی سیتوپلاسمی در حدود 60 تا 70 درصد پروتئین و 20 تا 30 درصد لیپید است که درآن مقدارزیادی فسفاتید وجود دارد و مقدارمواد قندی آن کم است. درغشاء سیتوپلاسمی پروکاریوت ها استرول وجود ندارد.
در بسیاری از باکتری ها خصوصاً در باکتری های گرم مثبت، رشته های کیسه مانند از غشای سیتوپلاسمی به داخل سیتوپلاسم می رود که مزوزوم نام دارد.
اعمال غشاء سیتوپلاسمی عبارتند از:
1- قابلیت نفوذ انتخابی : درانتقال مواد به داخل سلول که ممکن است همراه با مصرف انرژی (انتقال فعال) یا بدون مصرف انرژی (انتقال غیر فعال) باشد.
2- جایگاه واکنش های تنفسی : آنزیم های تنفسی (خصوصاً فسفریلاسیون اکسیداتیو) و سیتوکروم های مختلف (در زنجیره های انتقال الکترون) در داخل غشاء سیتوپلاسمی (بخصوص مزوزوم ها) واقعند.
3- دفع اگزوآنزیم ها و اگزوتوکسین ها : مولکول های کوچک مانند اسیدهای آمینه، نوکلئوزیدها و بعضی قندها نظیرگلوکز از غشاء وارد سلول می شوند. جذب این مواد ممکن است بصورت فعال (اکتیو) یا غیر فعال (پاسیو) انجام گیرد. اما ماکرو مولکول ها نمی توانند به راحتی از غشاء بگذرند. باکتری به منظور تبدیل ماکرومولکول ها به مولکول های ساده ی قابل جذب، اگزوآنزیم ها را ترشح می کند (در جانوران عالی این آنزیم ها به درون روده ریخته می شود). پروته آز، لیپاز، پلی ساکاریداز، آلکالن فسفاتاز و پنی سیلیناز نمونه هایی از اگزوآنزیم هاست.
4- جایگاه واکنش های بیوسنتز: غشای سیتوپلاسمی درعمل تقسیم سلولی نقش مهمی به عهده دارد و کلیه آنزیم ها و مولکول های ضروری برای بیوسنتز دیواره ی سلولی، لیپیدهای غشاء، قسمت های خارج سلول و همانند سازی DNA درغشاءسیتوپلاسمی واقعند(طبق شواهد امر در مزوزوم هایی که DNA به آن ها متصل است.)
3.دیواره سلولی (Cell wall):
دیواره سلولی، پروتوپلاست (شامل سیتوپلاست و غشای سیتوپلاسمی) را احاطه می کند و بلافاصله در قسمت خارجی پرده سیتوپلاسمی قرار دارد و این پرده را در مقابل فشار اسمزی زیاد داخلی (5 تا 25 اتمسفر)، حمایت می کند. شکل باکتری مربوط به سختی این لایه است و آنتی ژن پیکره ی میکروبی نیز در این قسمت قرار دارد.
ساختمان شیمیایی دیواره سلولی
ترکیب شیمیایی دیواره سلولی در انواع مختلف متفاوت است. ولی در تمام باکتری ها دارای یک ساختمان اصلی است که از پپتیدوگلیکان تشکیل شده است. علاوه بر پپتیدوگلیکان، دیواره سلولی دارای ترکیبات دیگری به نام ساختمان مخصوص نیز می باشد. پپتیدوگلیکان یک ساختمان مشبک، پیچیده و محکم است. مقاومت دیواره باکتری ها به دلیل همین ساختار مشبک است.
فقدان مجموعه ی پپتیدوگلیکان در سلول های پستانداران، استفاده ی بالینی مهمی دارد. به طوری که پنی سیلین و آنتی بیوتیک های دیگر، از ساخته شدن پپتیدوگلیکان جلوگیری و به این وسیله از تشکیل دیواره سلولی باکتری ها ممانعت می کنند. باکتری ها بر مبنای میزان پپتیدوگلیکان و نوع ساختمان مخصوص به دو گروه گرم مثبت(Gram positive) و گرم منفی (Gram negative) تقسیم می شوند.
ساختمان مخصوص دیواره سلولی
الف- ساختمان مخصوص دیواره سلولی باکتری های گرم مثبت:
در دیواره سلولی باکتری های گرم مثبت علاوه بر پپتیدوگلیکان، ساختمان مخصوصی نیز وجود دارد که شامل مقادیری از اسیدهای تئی کوئیک است. در بعضی از باکتری های گرم مثبت، ساختمان مخصوص ممکن است پلی ساکاریدی باشد. اسیدهای تئی کوئیک، پلی مرهای فسفاتی هستند که محتوی ریبیتول و یا گلیسرول می باشند و به وسیله ی پیوندهای دی استر، بهم متصل شده اند. اسید تئی کوئیک، بیشتر در بین لایه های پپتیدوگلیکان قرار داشته و احتمالا از منافذ لایه ی موکوپپتید، به خارج راه پیدا کرده است. اسید تئی کوئیک، آنتی ژن های مهم سطحی باکتری های گرم مثبت را تشکیل می دهد. علاوه بر این، مشخص کننده ی اختلاف سرولوژیکی گروه های مختلف در باکتری های گرم مثبت است.
ب- ساختمان مخصوص دیواره سلولی باکتریهای گرم منفی:
قسمت اعظم دیواره سلولی باکتری های گرم منفی را ساختمان مخصوص تشکیل می دهد. این ساختمان از لحاظ فیزیکی و شیمیایی به مراتب پیچیده تر از باکتری های گرم مثبت است و از لیپید، پروتئین و پلی ساکارید ساخته شده است. در ساختمان مخصوص گرم منفی ها، اسیدهای تئی کوئیک وجود ندارند. ساختمان مخصوص در این باکتری ها شامل سه لایه است که در سطح خارجی لایه ی پپتیدوگلیکان قرار دارند.
این لایه ها عبارتند از:
1-لایه ی لیپوپروتئین: پپتیدوگلیکان را به غشاء خارجی متصل می کند.
2-غشاء خارجی (Outer Membrane): ساختمان آن شبیه غشاء سیتوپلاسمی از دو لایه فسفولیپید است که لایه ی خارجی آن به وسیله ی مولکول های لیپوپلی ساکارید اشغال شده است. در غشاء خارجی، همانند غشاء سیتوپلاسمی، یک سری از مولکول های پروتئینی در ماتریکس فسفولیپید فرو رفته اند. علاوه بر این، نوع ویژه ای از پروتئین های منفذ دار به نام پورین هم در غشای خارجی یافت می شوند. پورین ها، پروتئین های مهمی هستند که تنها در غشاء خارجی وجود دارند. این پروتئین ها در ساخت منافذ یا کانال های غشایی شرکت می کنند.
3- لیپوپلی ساکارید (LPS): لیپوپلی ساکارید توسط پیوندهای هیدروفوبیک به غشاء خارجی متصل شده است. لیپوپلی ساکارید از دو بخش لیپید A و پلی ساکارید تشکیل شده است. "لیپید A" یک دی ساکارید با چند دم اسید چرب متصل به غشا است. لیپید A برای انسان سمی است و اندو توکسین باکتری های گرم منفی ها نام دارد . هنگامی که سلول های باکتریایی توسط کارکرد سیستم ایمنی بدن ما لیز می شود، قطعات غشای خارجی حاوی لیپید A به گردش خون آزاد می شود و باعث ایجاد تب، اسهال و حتی شوک آندوتوکسین کشنده می گردد که همچنین شوک سپتیک گفته می شود .
بخش پلی ساکاریدی لایه ی لیپوپلی ساکارید (LPS) از دو قسمت تشکیل شده است:
* قسمتی موسوم به هسته که در تمام باکتری ها یکسان است.
* واحدهای انتهایی تکرار شونده که در انواع مختلف باکتری ها متفاوت است.
|
شکل 4-1) ساختمان مولکولی لیپوپلی ساکارید |
واحدهای تکرارشونده معمولا تری، تتراد و پنتا ساکارید هستند. پلی ساکارید نقش آنتی ژنی مهمی را داراست که در باکتری های گرم منفی به آنتی ژن «O» موسوم است. ویژگی آنتی ژنیک، مربوط به واحدهای تکرار شونده ی انتهایی است که به صورت مولکول هایی در سطح خارجی سلول قرار گرفته و بر اساس اختلاف این واحدهای تکرار شونده، تقسیم بندی باکتری ها بر اساس گروه های آنتی ژنیکی صورت می گیرد. LPS دارای اثر فوق العاده سمی بر روی جانوران است و به آندوتوکسین باکتری های گرم منفی موسوم است. خاصیت سمی بودن LPS مربوط به بخش لیپید A آن است. این سم به آن سبب آندوتوکسین نامیده می شود که به سطح سلول متصل است و فقط پس از لیز باکتری آزاد می شود.
لیستریا که یک باسیل گرم مثبت است به طور استثنایی در دیواره سلولی خود اندوتوکسین دارد واین بسیار حیرت انگیزاست، زیرا تمام دیگر ارگانیسم های اندوتوکسین دار گرم منفی اند .

شکل 5-1) ساختمان جدار سلولی در باکتری های گرم مثبت و گرم منفی
اسفروپلاست (Spheroplast):
گاهی دیواره سلولی باکتری ضعیف می شود یا قسمتی از آن از بین می رود، برای مثال زمانی که باکتری در مجاورت با یک آنتی بیوتیک مهار کننده ساخت دیواره قرار می گیرد. در چنین حالتی باکتری را اسفروپلاست می نامند. اسفروپلاست به آسانی از باکتری های گرم منفی، هنگامی که در حضور ماده ای مانند پنی سیلین، سیکلوسرین یا گلایسین کشت داده شوند به دست می آید. این مواد از سنتز پپتیدوگلیکان دیواره سلولی جلوگیری می نمایند. فعالیت متابولیکی اسفروپلاست ها و توانایی تکثیر و بیماری زایی آنها در مقایسه با باکتری های دارای دیواره سالم بسیار کاهش می یابد. در عین حال اسفروپلاست ها قدرت بازگشت دارند و هنگامی که در یک محیط فاقد مهار کننده ی سنتز دیواره سلولی کشت داده شوند، تبدیل به سلول طبیعی می گردند. این موضوع اهمیت بالینی زیادی در زمینه ی عود یا بازگشت مجدد بعضی عفونت ها زمانی که برای درمان آنها به شکل ناقص از آنتی بیوتیکهای موثر بر دیواره استفاده می شود دارد. زمانی که یک باکتری دیواره خود را به طور کامل از دست می دهد به فرم پروتوپلاست تبدیل می گردد. چنین پروتوپلاست هایی دیگر امکان بازگشت به شکل یک سلول طبیعی دیواره دار را ندارند و با توجه به فشار اسمزی زیاد تر درون سلول نسبت به محیط اطراف باکتری، بیش از حد آب جذب می کنند، متورم می شوند و می ترکند.
کپسول و گلیکوکالیکس (Capsule & Gylcocalyx). بسیاری از باکتری ها وقتی در محیط طبیعی رشد می کنند مقادیر زیادی پلیمر خارج سلولی می سازند که بیشتر از نوع پلی ساکارید است. این لایه اگر متراکم و با حدود مشخص باشد کپسول و اگر به صورت یک شبکه شل از فیبریل ها به طرف خارج سلول امتداد داشته باشد گلیکوکالیکس نامیده می شود. کپسول در افزایش قدرت تهاجم باکتری های بیماری زا نقش دارد و غالبا آنها را از فاگوسیته شدن مصون می دارد. بعلاوه کپسول برای میزبان دارای خاصیت آنتی ژنی است و معمولا از آن با عنوان آنتی ژن K یاد می شود.گلیکوکالیکس نقش زیادی در چسبیدن و اتصال باکتری ها بطور غیر اختصاصی به سطح سلول های میزبان و یا به سطوح محیطی و یا به یکدیگر دارد؛ به همین دلیل گلیکوکالیکس باکتری های سطح دندان در تشکیل پلاک دارای نقش اساسی است.
دو تست مهم تشخیص کپسول یک کارشناس آزمایشگاه را قادر می سازد تا کپسول ها را در زیر میکروسکوپ ببیند و این به تشخیص نوع باکتری کمک می نماید:
رنگ آمیزی با مرکب هندی : (India ink stain) از آنجایی که مرکب هندی توسط کپسول جذب نمی شود، کپسول به شکل یک هاله شفاف در اطراف سلول دیده می شود. از این تست در تشخیص اولیه ی قارچ کریپتوکوکوس و نیز پنوموکوک کپسول دار استفاده می شود.
واکنش کوآلانگ (Quellung reaction): در این واکنش (که از آن به عنوان یک تست آزمایشگاهی در تشخیص باکتری های کپسول دار استفاده می شود) باکتری های کپسول دار با آنتی بادی های قابل اتصال به کپسول مخلوط می شوند. وقتی این آنتی بادی ها به کپسول باکتری متصل شوند کپسول ها متورم می گردند وتوسط میکروسکوپ قابل مشاهده اند.
آنتی بادی های ضد کپسول های باکتریایی از طریق تسهیل اتصال نوتروفیل ها و ماکروفاژها به باکتری های کپسول دار و بلعیدن آنها به بدن کمک می کنند. روند اتصال آنتی بادی ها به کپسول اپسونیزاسیون (opsonization) نام دارد. زمانی که اپسونیزاسیون انجام شود، ماکروفاژ یا نوتروفیل سپس می تواند به بخش Fc آنتی بادی اتصال یابد و باکتری را در بر گیرد (شکل 6-1).
یکی از واکسن ها ی ساخته شده علیه استرپتوکوکوس پنومونیه محتوی 23 آنتی ژن از شایع ترین انواع کپسول های این باکتری می باشد. ایمونیزاسیون با این واکسن باعث برانگیختن پاسخ ایمنی علیه آنتی ژن های کپسولی و تولید آنتی بادی می شود که فرد را در مقابل عفونت های آتی با باکتری مذکور مصون می سازد.

شکل 1-6: واکنش کوآلانگ
ضمایم جدار سلولی :
1. تاژک(Flagella) . باکتری های متحرک دارای زوایدی رشته مانند به نام تاژک هستند. این اندامک بصورت یک رشته از جنس پروتئینی به نام فلاژلین است. تاژک نازک ، موج دار و با قابلیت انعطاف است و باعث حرکت باکتری می گردد. ضخامت آن حدود 10 تا 30 نانومتر و طول آن معمولا چندین برابر طول باکتری است که گاهی به 80 میکرومتر نیز می رسد. تاژک توسط یک جسم پایه به باکتری متصل می شود. جسم پایه در تمام عمق دیواره سلول ادامه دارد و به غشای بیرونی و غشای سیتوپلاسمی در باکتری های گرم منفی و غشای سیتوپلاسمی در باکتری های گرم مثبت اتصال می یابد (باکتری های گرم مثبت غشای خارجی ندارند). جسم پایه به اطراف می چرخد و تاژک را به حرکت در می آورد (شکل 6-1). این سبب می شود تا تاژک با حرکات منظم موج دار خود باکتری را به طرف تراکم مواد شیمیایی نزدیک و یا از آن دور کند. این نوع حرکت باکتری کیمیوتاکسی) (Chemotaxis گفته می شود.
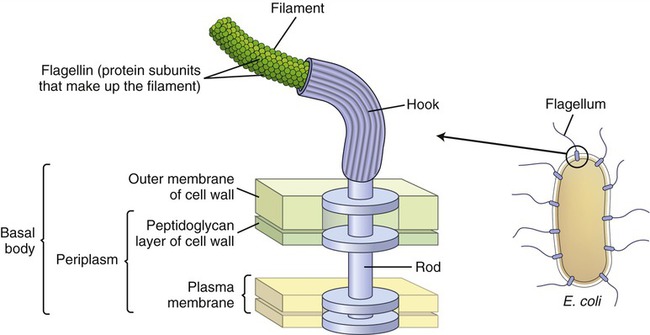
شکل 7-1) ساختمان داخلی یک تاژک
تاژک باکتری برای میزبان خاصیت آنتی ژنیک هم دارد و به نام آنتی ژن H نامیده می شود. آرایش تاژک یا تاژک ها در اطراف باکتری ها متفاوت است. تاژک ممکن است به تعداد یکی و فقط در یک قطب (تاژک مونوتریکوس)، یکی در هر قطب (تاژک آمفی تریکوس)، چند تا در هر قطب (تاژک لوفوتریکوس) و یا در سرتاسر پیرامون باکتری (تاژک پری تریکوس) وجود داشته باشد (شکل 7-1). برای مثال باکتری ویبریو کلرا دارای تاژک مونوتریکوس و باکتری های اشریشیا کولی و پروتئوس میرابیلیس دارای تاژک های پری تریکوس هستند.
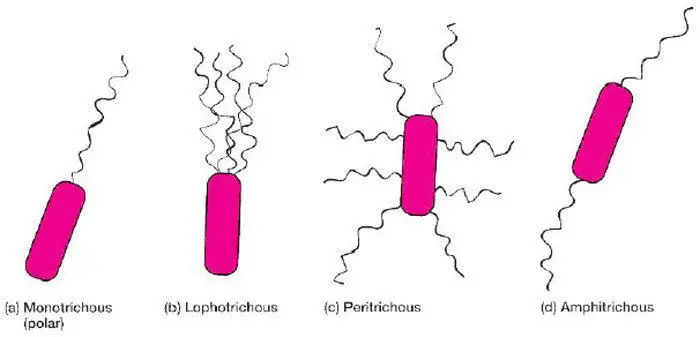
شکل 7-1) انواع آرایش تاژک در باکتری ها
2. پیلی یا فیمبریه(Pilli) . در سطح بسیاری از باکتری های گرم منفی و برخی از از باکتری های گرم مثبت کرک های ظریفی به قطر 5 تا 10 نانومتر و طول تقریبی 1 تا 10 میکرومتر وجود دارد که سطح باکتری را پوشانده اند. تعداد آنها در سطح باکتری بین 10 تا 500 عدد و یا بیشتر است. پیلی کوتاهتر و نازک تر از تاژک است. بعلاوه کم و بیش مستقیم و بدون قابلیت انعطاف است. پیلی از نوعی پروتئین به نام پیلین ساخته شده و دارای خاصیت آنتی ژنیک است. پیلی عامل چسبندگی باکتری به سطح سلول های میزبان است. این اتصال از طریق اتصال پیلی به گیرنده های اختصاصی در سطح سلول صورت می گیرد و در این صورت به پیلی ادهزین (Adhesin) گفته می شود. بسیاری از باکتری های بیماری زا دارای ادهزین هایی هستند که برای توانایی آنها در ایجاد بیماری ضروری هستند. برای مثال نایسریا گونوره آ دارای پیلی هایی است که به باکتری امکان اتصال به سلول های گردن رحم و سلول های buccal و ایجاد بیماری سوزاک را می دهد .باکتری های اشریشیا کولی و کمپیلوباکتر ژژونی بدون اتصال ادهزین های خود به اپیتلیوم روده نمی توانند باعث اسهال شوند. باکتری بوردتلا پرتوسیس نیز از ادهزین های خود به منظور اتصال به سلول های مژه دار تنفسی و ایجاد سیاه سرفه استفاده می کند. باکتری های فاقد پیلی معمولا امکان اتصال اختصاصی به سطوح میزبانی، کلونیزاسیون و بیماریزایی را ندارند.
پیلی جنسی(Sex pilli) . این نوع پیلی از انواع دیگر پیلی بلند تر و ضخیم تر است و فقط در باکتری های دهنده ی ماده ژنتیکی (باکتری F+) مشاهده می شود. تعداد آنها در هر باکتری کم و فقط به تعداد 1تا 4 عدد است. این نوع پیلی به هنگام پدیده کونژوگاسیون در انتقال DNA از باکتری دهنده (F+) به باکتری گیرنده (F-) نقش دارد.
شکل 8-1. پیلی جنسی
اسپور و اسپورزایی (Spore & Sporulation) :
اکثر باکتری ها فقط دارای زندگی رویشی هستند. در این نوع زندگی باکتری دارای فعالیتهای حیاتی آشکار از قبیل رشد و نمو، تولید اگزوتوکسین و اگزوانزیم، حرکت ، متابولیسم مشخص و ... می باشد. شکل رویشی باکتری به شرایط نامساعد محیطی نسبتا حساس است. گروهی از باکتری ها قادرند در شرایط نامساعد محیطی به مرحله ای از زندگی وارد شوند که آنها را نسبتا مقاوم می کند. این شکل از باکتری از لحاظ ساختمان و عمل دقیقا برعکس شکل رویشی باکتری است و اسپور نام دارد. اسپور از نظر متابولیکی شکل خفته باکتری است و فقط در چرخه زندگی برخی از باکتری ها دیده می شود.
هنگام اسپورسازی تغییرات بیوشیمیایی پیچیده و شگرفی در سلول باکتری بوجود می اید. بطوریکه لایه های ضخیمی دور تا دور بخشی از سلول باکتری را فرا می گیرد. باکتری درحال اسپورسازی قسمت اعظم آب و آنزیم های مرحله رویشی خود را از دست می دهد و وارد مرحله زندگی غیر فعال یا نهفته می شود. در چنین شرایطی فعالیت های متابولیکی باکتری بسیار کند می شود و هیچ گونه فعالیت حیاتی آشکاری را نشان نمی دهد. اسپور می تواند شرایطی از قبیل حرارت، سرما، خشکی، مواد شیمیایی و حتی پرتوها را تا حد زیادی تحمل کند. در شرایط مساعد اسپور مجددا طی مراحلی تندش می کند و از آن یک سلول رویشی حاصل می گردد. خانواده باسیلاسه و کلستریدیوم مهم ترین باکتری های بیماری زایی هستند که قادر به تولید اسپورند.
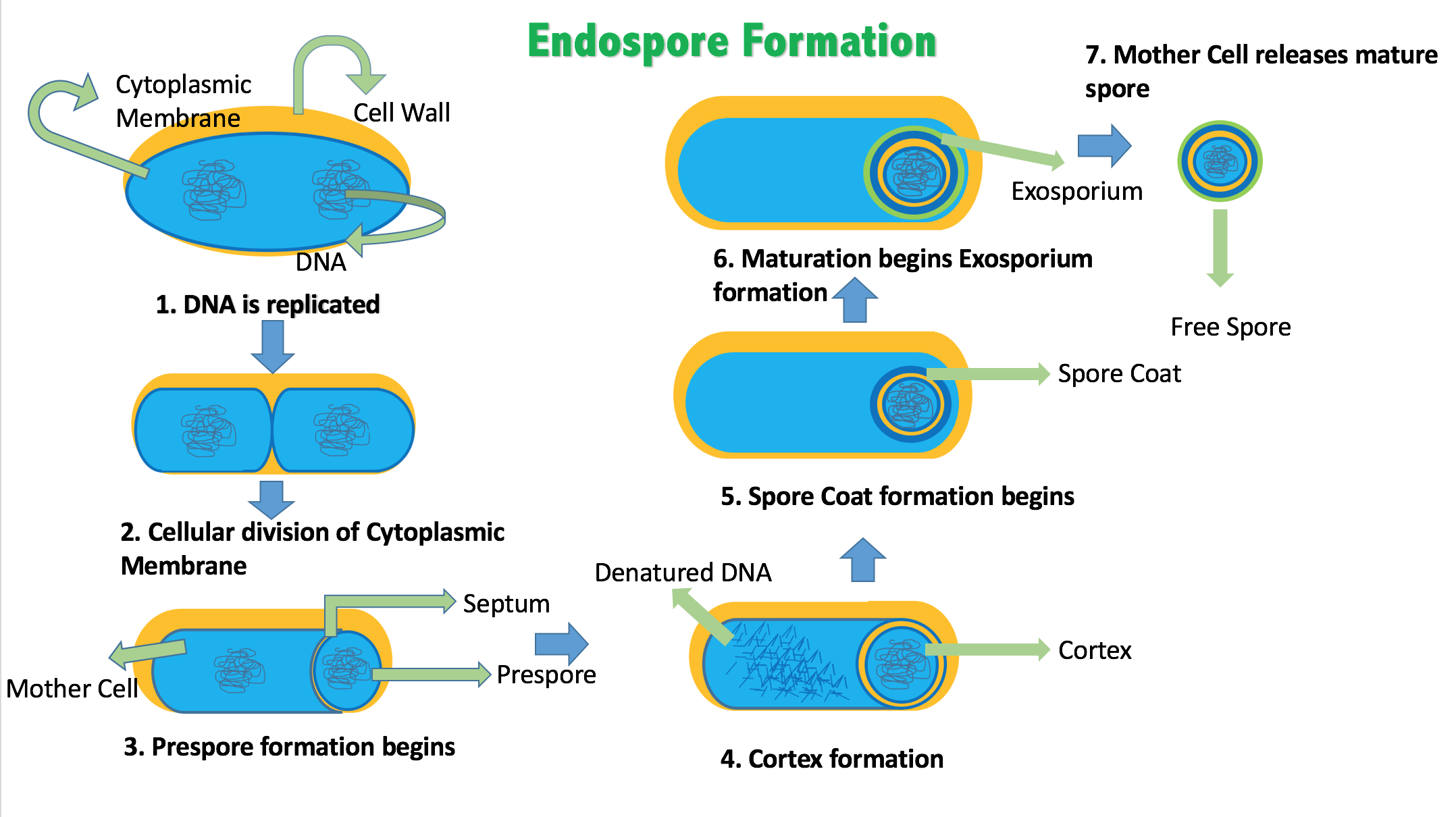
شکل 9-1. مراحل اسپور سازی در یک باکتری اسپور زا

